La glucólisis, del griego glycos (azúcar) y lysis (ruptura, destrucción, transformación), es la ruta metabólica encargada de oxidar la glucosa con la finalidad de obtener energía para la célula. Consiste en 10 reacciones enzimáticas consecutivas que convierten a la glucosa en dos moléculas de piruvato, el cual es capaz de seguir otras vías metabólicas y así continuar entregando energía al organismo.
El tipo de glucólisis más común y más conocida es la vía de Embden-Meyerhof, explicada inicialmente por Gustav Embden y Otto Fritz Meyerhof. No obstante, el vocablo «glucólisis» se usa con frecuencia como sinónimo de la vía de Embden-Meyerhof.
Los primeros estudios informales de los procesos glucolíticos fueron iniciados en 1860, cuando Louis Pasteur descubrió que los microorganismos son los responsables de la fermentación, y en 1897 cuando Eduard Buchner encontró que cierto extracto celular puede causar fermentación. La siguiente gran contribución fue de Arthur Harden y William Young en 1905, quienes determinaron que para que la fermentación tenga lugar son necesarias una fracción celular de masa molecular elevada y termosensible (enzimas) y una fracción citoplasmática de baja masa molecular y termorresistente (ATP, ADP, NAD+ y otras coenzimas). Los detalles de la vía en sí se determinaron en 1940, con un gran avance de Otto Meyerhoff y algunos años después por Luis Leloir.
En eucariotas y procariotas, la glucólisis ocurre en el citosol de la célula. En células vegetales, algunas de las reacciones glucolíticas se encuentran también en el ciclo de Calvin, que ocurre dentro de los cloroplastos.
Fases de la Glucólisis
La glucólisis es la forma más rápida de conseguir energía para una célula y, en el metabolismo de carbohidratos, generalmente es la primera vía a la cual se recurre. Durante la glucólisis se obtiene un rendimiento neto de dos moléculas de ATP; el ATP puede ser usado como fuente de energía para realizar trabajo metabólico, mientras que el NADH puede tener diferentes destinos.
La glucólisis se divide en dos fases principales:
-
Fase de Gasto de Energía: La primera fase consiste en transformar una molécula de glucosa en dos moléculas de gliceraldehído (una molécula de baja energía) mediante el uso de 2 ATP.
-
Paso 1: Fosforilación de la glucosa. La primera reacción de la glucólisis es la fosforilación de la glucosa, para activarla y así poder utilizarla en otros procesos. Esta activación ocurre por la transferencia de un grupo fosfato del ATP, una reacción catalizada por la enzima hexoquinasa, la cual puede fosforilar (añadir un grupo fosfato) a moléculas similares a la glucosa, como la fructosa y manosa.
Las ventajas de fosforilar la glucosa son dos: la primera es hacer de la glucosa un metabolito más reactivo, y la segunda es que la glucosa-6-fosfato no puede cruzar la membrana celular -a diferencia de la glucosa-, ya que la carga negativa que le proporciona el grupo fosfato a la molécula hace que sea más difícil atravesarla. De esta forma se evita la pérdida de sustrato energético para la célula. En numerosas bacterias esta reacción está acoplada a la última reacción de la glucólisis (de fosfoenolpiruvato a piruvato) para poder aprovechar la energía sobrante de la reacción.
-
Paso 2: Isomerización de la glucosa-6-fosfato. En esta reacción, la glucosa-6-fosfato se isomeriza a fructosa-6-fosfato, mediante la enzima glucosa-6-fosfato isomerasa. Este es un paso importante, puesto que aquí se define la geometría molecular que afectará los dos pasos críticos en la glucólisis: El próximo paso, que agregará un grupo fosfato al producto de esta reacción, y el paso 4, cuando se creen dos moléculas de gliceraldehido que finalmente serán las precursoras del piruvato.
-
Paso 3: Fosforilación de la fructosa-6-fosfato. Fosforilación de la fructosa 6-fosfato en el carbono 1, con gasto de un ATP, a través de la enzima fosfofructoquinasa-1 (PFK1). Este fosfato tendrá una baja energía de hidrólisis. Por el mismo motivo que en la primera reacción, el proceso es irreversible. El nuevo producto se denominará fructosa-1,6-bisfosfato.
La irreversibilidad es importante, ya que la hace ser el punto de control de la glucólisis. Como hay otros sustratos aparte de la glucosa que entran en la glucólisis, el punto de control no está colocado en la primera reacción, sino en esta.
-
Paso 4: Escisión de la fructosa-1,6-bisfosfato. La enzima aldolasa (fructosa-1,6-bisfosfato aldolasa), mediante una condensación aldólica reversible, rompe la fructosa-1,6-bisfosfato en dos moléculas de tres carbonos (triosas): dihidroxiacetona fosfato y gliceraldehído-3-fosfato.
-
Paso 5: Isomerización de la dihidroxiacetona fosfato. Puesto que solo el gliceraldehído-3-fosfato puede seguir los pasos restantes de la glucólisis, la otra molécula generada por la reacción anterior (dihidroxiacetona-fosfato) es isomerizada (convertida) en gliceraldehído-3-fosfato. Este es el último paso de la "fase de gasto de energía". Solo se ha consumido ATP en el primer paso (hexoquinasa) y el tercer paso (fosfofructoquinasa-1). Cabe recordar que el 4.º paso (aldolasa) genera una molécula de gliceraldehído-3-fosfato, mientras que el 5.º paso genera una segunda molécula de este.
-
-
Fase de Producción de Energía: En la segunda fase, el gliceraldehído se transforma en un compuesto de alta energía, cuya hidrólisis genera una molécula de ATP, y como se generaron 2 moléculas de gliceraldehído, se obtienen en realidad dos moléculas de ATP. Esta obtención de energía se logra mediante el acoplamiento de una reacción fuertemente exergónica después de una levemente endergónica. Este acoplamiento ocurre una vez más en esta fase, generando dos moléculas de piruvato.
-
Paso 6: Oxidación del gliceraldehído-3-fosfato. Técnicamente, el grupo aldehído se oxida a un grupo acil-fosfato, que es un derivado de un carboxilo fosfatado. Mientras el grupo aldehído se oxida, el NAD+ se reduce, lo que hace de esta reacción una reacción redox.
-
Paso 7: Formación de ATP. En este paso, la enzima fosfoglicerato quinasa transfiere el grupo fosfato de 1,3-bisfosfoglicerato a una molécula de ADP, generando así la primera molécula de ATP de la vía. Como la glucosa se transformó en 2 moléculas de gliceraldehído, en total se recuperan 2 ATP en esta etapa. Los pasos 6 y 7 de la glucólisis nos muestran un caso de acoplamiento de reacciones, donde una reacción energéticamente desfavorable (paso 6) es seguida por una reacción muy favorable energéticamente (paso 7) que induce la primera reacción.
-
Paso 8: Isomerización del 3-fosfoglicerato. Se isomeriza el 3-fosfoglicerato procedente de la reacción anterior dando 2-fosfoglicerato, la enzima que cataliza esta reacción es la fosfoglicerato mutasa. Lo único que ocurre aquí es el cambio de posición del fosfato del C3 al C2.
-
Paso 9: Deshidratación del 2-fosfoglicerato. La enzima enolasa propicia la formación de un doble enlace en el 2-fosfoglicerato, eliminando una molécula de agua formada por el hidrógeno del C2 y el OH del C3.
-
Paso 10: Formación de piruvato y ATP. Desfosforilación del fosfoenolpiruvato, obteniéndose piruvato y ATP. Reacción irreversible mediada por la piruvato quinasa. El enzima piruvato quinasa es dependiente de magnesio y potasio.
-
Regulación alostérica y hormonal de la glucólisis
Regulación de la Glucólisis
La fosfofructoquinasa-1 es la enzima principal de la regulación de la glucólisis. Actúa como una llave de agua: si está activa cataliza muchas reacciones y se obtiene más fructosa-1,6-bisfosfato, lo que permitirá a las enzimas siguientes transformar mucho piruvato. Si está inhibida, se obtienen bajas concentraciones de producto y por lo tanto se obtiene poco piruvato. La fosfofructoquinasa tiene centros alostéricos, sensibles a las concentraciones de intermediarios como citrato y ácidos grasos.
La hexoquinasa es un punto de regulación poco importante, ya que se inhibe cuando hay mucho G-6P en músculo.
El rendimiento total de la glucólisis de una sola glucosa (6C) es de 2 ATP y no 4 (dos por cada gliceraldehído-3-fosfato (3C)), ya que se consumen 2 ATP en la primera fase, y 2 NADH (que dejarán los electrones Nc en la cadena de transporte de electrones para formar 3 ATP por cada electrón).

Deficiencia de Fosfofructoquinasa-1 (PFK-1)
La deficiencia de fosfofructoquinasa-1 (PFK-1) es una forma poco frecuente de enfermedad por almacenamiento de glucógeno (EAG) que se caracteriza clásicamente por fatiga de esfuerzo e intolerancia al ejercicio muscular.
Signos Clínicos y Diagnóstico
Los signos clínicos incluyen la intolerancia al ejercicio muscular (a menudo con rabdomiólisis) con calambres musculares y dolor durante el ejercicio. Los pacientes no presentan un fenómeno de recuperación muscular y los síntomas pueden empeorar tras la ingesta de glucosa. La confirmación diagnóstica no invasiva se realiza mediante el análisis molecular del gen PFKM. Cuando se realiza, la biopsia muscular suele mostrar miopatía vacuolar, aumento del contenido de glucógeno y deficiencia de la enzima PFK.
Genética y Asesoramiento
La transmisión es autosómica recesiva. Debe ofrecerse asesoramiento genético a las parejas en riesgo (aquellas en las que ambos individuos son portadores de una mutación causante de la enfermedad) e informar de que la probabilidad de tener un descendiente afecto es del 25% en cada embarazo. Los episodios de rabdomiólisis pueden prevenirse evitando el ejercicio intenso.
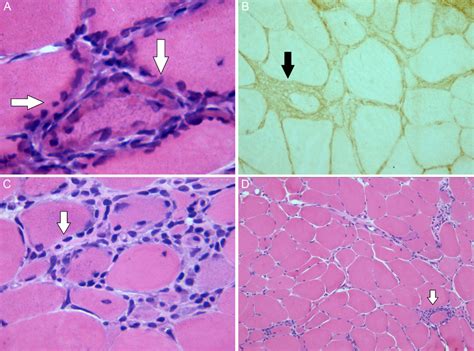
Impacto en la Fermentación Láctica
La deficiencia de PFK-1 afecta directamente la capacidad de la célula para realizar la glucólisis de manera eficiente, lo cual tiene un impacto significativo en la fermentación láctica. Al ser la PFK-1 la principal enzima reguladora de la glucólisis, su deficiencia provoca una disminución en la producción de fructosa-1,6-bisfosfato y, consecuentemente, en la formación de piruvato. El piruvato es el sustrato clave para la fermentación láctica, un proceso anaeróbico que permite la regeneración del NAD+ necesario para que la glucólisis continúe en ausencia de oxígeno.
En individuos con deficiencia de PFK-1, la producción reducida de piruvato limita la capacidad de los músculos para generar ATP rápidamente durante el ejercicio intenso a través de la glucólisis anaeróbica. Esto conduce a una acumulación de metabolitos aguas arriba del bloqueo enzimático y una menor producción de lactato, lo que contribuye a los síntomas de fatiga e intolerancia al ejercicio muscular.
tags: #deficit #en #fosfofructoquinasa #1 #fermentacion #lactica